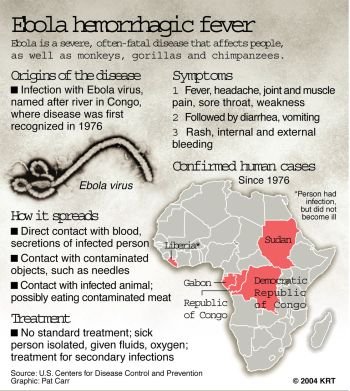
Ebola death in Mali sparks panic (VIDEO)
Mali Becomes 6th African Nation with Ebola, Despite WHO’s Confidence! (VIDEO)
Ebola Spreads to Mali as WHO Races to Contain Outbreak
As Ebola Spreads, Asia Senses Vulnerability (NYT)
Ebola Website – World Health Organization
Faecal–oral transmission
Viruses that target the gut are just as diverse as respiratory viruses and, in the same way, the hundreds of different gut virus types can attack throughout life. These viruses are spread either directly by unwashed hands or via drinking water, food, and contaminated objects like surfaces and blankets; they are also highly adapted to our bodies and our lifestyle. They survive the acid environment of the stomach that kills most other invaders and then attack the gut lining, killing the cells and thereby stopping the production of digestive enzymes and preventing fluid absorption. All this induces the unpleasant symptoms of gastroenteritis. These viruses manufacture huge numbers of offspring that can survive for long periods outside the body, and infect with a very low virus dose. After an incubation period of between one and two days, the two key culprits, rotaviruses and noroviruses, induce sudden onset of projectile vomiting, profuse watery diarrhoea, and abdominal cramps, which effectively contaminate the environment and ensure their own survival.
Rotaviruses are a major cause of gastroenteritis globally, particularly targeting children under 5. The disease varies in severity but usually lasts four to seven days, with the main problem being dehydration. Indeed, rotaviruses cause over 600,000 infant deaths a year worldwide, mostly in developing countries where the viruses spread easily and emergency rehydration procedures are not always available. With up to a hundred billion (10 11) virus particles in each milliliter of faeces produced by an infected child, and only 10 virus particles actually required to pass the infection on, it is not surprising that rotavirus outbreaks are frequent and difficult to control.
As they circulate in the community, rotaviruses, like flu viruses, undergo genetic drift, accumulating point mutations until they are sufficiently different to infect those already immune to the parent virus strain. Also, many strains of rotavirus cause gastroenteritis in young animals such as calves, piglets, lambs, foals, chickens, and rabbits, which can act as rotavirus reservoirs. Again, like flu viruses, from time to time human rotaviruses undergo a genetic shift by gene reassortment with animal rotaviruses.
This can produce an entirely new strain with the potential to cause a widespread epidemic. Noroviruses are the second most common cause of viral gastroenteritis after rotaviruses, producing a milder disease of shorter duration. These viruses account for around 23 million cases of gastroenteritis every year, with epidemics commonly centering on nursing homes, hospitals, and children’s nurseries, camps, and schools. Unusually, immunological memory to noroviruses tends to be short, so epidemics affect adults as well as children.
Outbreaks among the passengers and staff of cruise ships often hit the headlines, not only ruining the luxury holiday for those on board but also causing severe loss of revenue for the cruise company as ships often have to be taken out of service while the source of the outbreak is identified and the ship disinfected.
The comparative sizes of a typical bacterium and representative viruses al (t example, a cruise ship bound for Alaska set sail from Vancouver with 1,218 passengers and 564 crew on board. The very next day, 5 passengers came down with gastroenteritis, and by the time they disembarked 7 days later, a total of 176 people had reported sick. In port, the ship was disinfected before taking on another group of holidaymakers, this time 1,336 passengers and 571 crew. On this trip, 219 people developed gastroenteritis, necessitating cancellation of the next cruise while the ship was subject to ‘aggressive cleaning and sanitizing’.
The environmental health inspectors could find no source of infection or ‘sanitary deficiencies’. This is commonly the case, and it just goes to show what an effective spreading strategy these viruses have evolved. The virus induces projectile vomiting, a single episode of which releases up to three million virus particles, theoretically enough to infect 300,000 people.
Enteroviruses are an unusual group of viruses because although, as the name suggests, they spread by the faecal-oral route, infect the gut, and are excreted in faeces, they only cause problems if they spread to other organs. Poliovirus is the best known in the group as it can cause a life-threatening disease, paralytic poliomyelitis, but only in around 1 in 1,000 of those it infects.
Like other enteric viruses, poliovirus can survive happily for long periods in water and sewage, so, where standards of hygiene are low, it spreads rapidly among young children. Polioviruses grow in the living cells of the gut and its associated lymph glands, producing no symptoms but, in a few cases, they target nervous tissues, where they may cause severe disease.
In the unlucky few, the virus homes to the brain, causing meningitis (called non-paralytic polio), or to the spinal cord, where it destroys nerve cells and paralyses the related muscles (paralytic polio). The latter is fatal in around 5% of cases, mainly when the paralysis involves the respiratory muscles, leading to respiratory failure.
Poliomyelitis is a disease of modern times, having risen to prominence in the West only in the 20th century. At one time, it caused terrifying summer outbreaks, seeming to strike indiscriminatingly at perfectly healthy children rather than spread from person to person. This was only halted when the vaccine was introduced in the 1960s.
In developing countries at this time and, it is assumed, before the 20th century in industrialized countries, polioviruses circulated freely in the community and infected virtually the whole population during early childhood. In this situation, paralytic poliomyelitis was almost unknown.
The silent nature of the infection is thought to result from residual maternal antibodies which passed across the placenta while the child was in utero, and protected it from paralytic disease by preventing viral spread outside the gut. Then, as standards of hygiene rose and infection in infancy became less common, many mothers remained uninfected and so had not generated antibodies that could be passed on to, and protect, their infants. Thus the incidence of paralytic polio was inversely related to levels of hygiene, rising along with industrialization of a nation.
Many virus families such as rotaviruses that rely on faecal-oral transmission and cause gastroenteritis in humans produce the same symptoms in animals, resulting in great economic loss to the farming industry. However, over the centuries, Rinderpest virus, the cause of cattle plague, has probably been responsible for more loss and hardship than any other.
Rinderpest virus is closely related to measles virus, but the disease it causes is very different. The virus infects cloven-hoofed animals such as oxen, buffalo, yak, sheep, goats, pigs, camels, and several wild species including hipD. H. Crawford, Rinderpest used to be a major problem in Europe and Asia, and when it was introduced into Africa in the late 19th century, it killed over 90% of cattle, with devastating economic loss. The Global Rinderpest Eradication Programme was set up in the 1980s aiming to use the effective vaccine to rid the world of the virus by 2010.
This was successful, and in October 2010 the disease was officially declared eradicated, the first animal disease and second infectious disease ever to be eliminated. Many acute infectious viruses thrive in hospital and care home settings, causing outbreaks of hospital acquired, or nosocomial, infections.
Although today’s headlines are generally dominated by notorious bacterial infections like MRSA (methicillin-resistant Staphylococcus aureus), Clostridium difficile, and the ‘flesh-eating bug’ Streptococcus pyogenes, nosocomial virus infections go unreported and are in fact a common cause of outbreaks severe enough to lead to ward closures. Unfortunately, in the close confines of a hospital ward, patients are easy prey for viruses. Viruses that circulate in the community causing silent or mild infections can be devastating to premature babies, those debilitated by cancer or other chronic illnesses, the elderly, and the immunosuppressed.
Most often, a recently admitted patient is the source of the infection but, not uncommonly, it is a staff member who may remain healthy and be totally unaware that he or she is spreading a potentially lethal virus. Norovirus, with its abrupt onset of projectile vomiting, is particularly difficult to control and, because its incubation period of 1 to 2 days is too short to allow identification of the source in time to prevent secondary spread.
Persistent viruses
Viruses fight a constant battle against host immunity, and for most there is just a small window of opportunity in which to reproduce and make a hasty exit before being wiped out by the formidable array of host defences. But some viruses have evolved strategies for overcoming these immune mechanisms and survive inside their host for prolonged periods, even for a lifetime.
Although the detailed mechanisms involved in these evasion strategies are very complex and varied, overall they encompass three basic manoeuvres: finding a niche in which to hide from immune attack, manipulating immune processes to benefit the virus, and outwitting immune defences by mutating rapidly.
Most persistent viruses have evolved to cause mild or even asymptomatic infections, since a life threatening disease would not only be detrimental to the host but also deprive the virus of its home. Indeed, some viruses apparently cause no ill effects at all, and have been discovered only by chance. One example is TTV, a tiny DNA virus found in 1997 during the search for the cause of hepatitis and named after the initials (TT) of the patient from whom it was first isolated.
We now know that TTV, and its relative TTV-like mini virus, represent a whole spectrum of similar viruses that are carried by almost all humans, non-human primates, and a variety of other vertebrates, but so far they have not been associated with any disease. With modern, highly sensitive molecular techniques for identifying non-pathogenic viruses, we can expect to find more of these silent passengers in the future.
The frequency with which viruses succeed in persisting in their hosts varies, with herpes viruses virtually always establishing a lifelong relationship that usually does no harm to the host.
Retroviruses also generally infect for life, but they may, like HIV, cause a disease in those they infect after a prolonged silent period. Other viruses, such as hepatitis B virus, struggle to evade the immune response, and many hosts eventually manage to clear the virus. Further, there are a few viruses that are usually cleared after primary infection but on rare occasions may stay put. Measles virus, for example, for unknown reasons persists after the acute infection in around 1 in 10,000 cases causing a fatal brain disease called sub acute sclerosing pan encephalitis (SSPE).
Because of the lifelong presence of foreign (viral) genes inside a host cell, a persistent virus can sometimes drive the cell it lodges in into uncontrolled growth, that is, to become cancerous. These include human T lymphotropic virus, hepatitis B and C viruses, Epstein–Barr virus, Kaposi sarcoma associated virus, and the papilloma viruses.
The herpesvirus family
Herpes viruses form an ancient family whose common ancestor probably evolved during the Devonian period around 400 million years ago when fish-like creatures were just emerging from the seas to inhabit dry land. In doing so, they must have encountered an array of ‘new’ microbes, among them the primitive phage-like viruses thought to be the ancestors of modern-day herpes viruses.
From this early beginning, herpes viruses have co-evolved with their hosts, each partner exerting selective pressure on the other until they have become remarkably well adapted to each other’s lifestyles, allowing the viruses to thrive long term, generally without detriment to the hosts. As their host species diverged, herpes viruses also diverged, so that now almost all species of mammals, birds, reptiles, amphibians, fish, and even some non-vertebrates, have their own particular herpes virus cocktail.
To date, over 150 different herpes viruses have been identified, all of which are large, enveloped DNA viruses coding for between 80 and 150 proteins. They are fragile viruses that cannot survive independently for long in the outside world, and so they tend to spread by close contact between infectious and susceptible hosts.
Without exception, herpes viruses establish a lifelong infection, often called a latent infection. The viruses survive inside host cells in a dormant state, having shut down their protein production and thereby having become invisible to host immunity. Occasionally, during the lifetime of the host this latent infection reactivates to produce new viruses. The evolution of this long-term strategy ensures that virus offspring reach a young and susceptible host population and thereby guarantees their survival.
There are three herpes virus subfamilies: alpha, beta, and gamma, with members categorized according to their biological properties, particularly the cell types in which they establish latency. So far, eight human herpes viruses have been discovered, named herpes virus (HHV) 1 to 8 in order of their discovery, but also given ‘common’ names by which they are more familiarly known.
We inherited these viruses from our primate ancestors, and so each has a counterpart in primates to which it is more closely related than it is to the other human herpes viruses. Having co-evolved with us, herpes viruses infect all human populations worldwide, including the most isolated Amerindian tribes.
It is generally assumed that in the past all the human herpes viruses were ubiquitous, but today their prevalence varies, the hierarchy perhaps reflecting their success at spreading between hosts in the modern world. Human herpes viruses can spread in a variety of ways: transmitted directly from mother to child in breast milk (CMV) or spread among family members and close contacts via saliva (HSV-1, CMV, EBV, HHV-6 and -7, KSHV). Of these viruses, HHV-6 and -7 are the most successful, infecting almost everyone worldwide.
The prevalence of EBV, HSV-1, and CMV is also high, but each has experienced a recent drop in areas where high standards of hygiene tend to block their transmission. Interestingly, HSV-2 and KSHV have a much lower prevalence than the other human herpes viruses and show a more restricted geographical distribution, being most common in parts of Africa.
These viruses rely on salivary transmission in childhood (KSHV) and/or sexual transmission between adults, and scientists speculate that they are the most vulnerable to recent cultural and lifestyle changes and therefore their worldwide distribution is the first to be significantly eroded.
The alpha human herpes viruses, HSV-1 and -2, are 85% identical at the DNA level, but traditionally HSV-1 causes a cold sore on the face whereas HSV-2 causes genital herpes. Although this is still generally true, in fact both viruses can infect the skin of the face and genital area, and a rising minority of genital herpes cases is now caused by HSV-1.
HSV-1 and -2 access the b to induce immunity without severeb37ody through a cut or abrasion and target skin cells where they replicate, killing the infected cells as new viruses are produced.
The majority of primary HSV infections are silent, but they sometimes cause a painful rash of tiny blisters in and around the mouth or in the genital area. With each blister containing thousands of virus particles; it is easy to see how the virus spreads to other individuals. HSV infection of the skin soon attracts the attention of immune cells and the lesions heal rapidly, but not before some virus particles have secretly infected nerve endings in the skin and climbed up the nerve fibers to the cell nucleus where they establish latency.
HSV from a facial infection (mainly HSV-1) goes latent in the trigeminal ganglia at the base of the skull, whereas viruses from genital lesions (mainly HSV-2) head for the sacral ganglia alongside the lower spinal column. As nerve cells survive for the life of the host and do not divide, they are an ideal site for a virus to lie low for a while.
But to assure its long-term survival, at some stage the virus must wake up and move on. So from time to time, new viruses are produced, which travel down the nerve fibres and are shed into saliva or genital secretions.
This reactivation may be silent or may manifest as a cold sore on the face, classically on or near the lips, in around 40% of those carrying HSV-1, and as genital herpes in around 60% of those carrying HSV-2. The triggers for HSV reactivation in an individual carrier are often quite clear and recognizable: decreased immunity due to drugs or illness, fever, increased levels of ultraviolet light (classically precipitated by a skiing trip), or menstruation and stress, but the molecular mechanisms involved are not understood.
Chickenpox, as a very common, acute infection of childhood, but being a herpes virus, VZV establishes a latent infection in virtually everyone it infects. Like the HSVs, VZV hides in nerve cells, but as the chickenpox rash is widespread on the body, the virus may lodge in the spinal ganglia related to any or all of the nerves supplying the skin.
Latent VZV can reactivate to cause shingles at any time in life, but this is most common in the elderly. Reactivation usually occurs in a single nerve cell, causing the typical painful shingles rash of tiny blisters along the course of that particular nerve. As infectious viruses are shed from these lesions, individuals who have not had it before can catch chickenpox from them. But shingles is not caught either from cases of shingles or chickenpox, as it is the result of reactivation of internal, latent viruses.
As with the HSVs, the molecular mechanisms involved in VZV reactivation are unknown, and why it should occur most commonly in nerves supplying the eye, neck, and trunk is also a mystery. However, again similar to HSV, reactivation is more common in patients with immunosuppression, including those who are HIV positive, have had an organ transplant, or are receiving chemotherapy. In all these groups, the rash may be severe, widespread, and even life-threatening, but several antiviral agents, including aciclovir, can have a beneficial effect.
Of the three human beta herpes viruses, CMV is the only one that causes significant health problems. Although the virus infects most people silently, it occasionally causes a glandular-fever-like illness at primary infection. But more importantly, the virus in a pregnant woman’s blood may on rare occasions cross the placenta and infect her unborn child. When this happens, it causes cytomegalic inclusion disease in around 10% of affected infants, inducing a wide range of symptoms including growth retardation, deafness, abnormalities of internal organsre0S blood clotting, and inflammation of the liver, lungs, heart, and brain.
CMV establishes latency in the bone marrow stem cells that develop into blood monocytes and tissue macrophages. These cells transport the latent virus via the blood to the tissues where virus reactivation is common. In healthy hosts, this is dealt with by the immune system without causing disease, but CMV replication produces significant pathology in immunosuppressed patients, and was responsible for blindness, severe diarrhoea, pneumonia, and encephalitis in many HIV-positive people before effective antivirals were developed in the early 1990s.
The two human gamma herpes viruses, EBV and KSHV, are both tumour viruses. However, although KSHV appears to cause no problems on primary infection, EBV may cause glandular fever, also called infectious mononucleosis. EBV generally infects silently during childhood, but if infection is delayed until adolescence or early adulthood, it causes glandular fever in around one-quarter of cases.
As childhood infection is virtually ubiquitous in developing countries, and is also very common in low socioeconomic groups in developed countries, glandular fever is most prevalent in high socioeconomic groups in the developed world. In these situations, it is quite common among senior school pupils and university students, estimated to affect around 1 in 1,000 university students per year in one UK study.
EBV infects and establishes latency in blood B cells, and perhaps because these cells are themselves part of the immune system, the infection engenders an exaggerated T cell response. Indeed, the symptoms of glandular fever, which typically include sore throat, fever, enlarged glands in the neck, and fatigue, are immune pathological in nature, caused by this massive outpouring of T cells rather than directly by the virus infection itself. Although the illness usually resolves over 10 to 14 days, fatigue may persist for up to 6 months, sometimes causing quite severe disruption to the sufferer’s way of life.
On rare occasions, EBV causes tumours and has also been suggested as the cause of several other diseases, particularly autoimmune diseases such as rheumatoid arthritis and multiple sclerosis.
The retrovirus family
Retroviruses infect a wide range of animal species, often acting as a silent passenger, but sometimes causing immunodeficiency, leukaemia, or solid tumours. There are several retroviruses that cause immunodeficiency in humans all of which have been acquired from primates. Today, these HIVs are the only non-tumour-forming retroviruses to cause disease in humans, but there are intriguing clues to suggest that ancient hominids may have been prey to several more. Evidence for this theory comes from the large number of identifiable retroviral remnants within the human genome, but how and when they got there, and why they have been retained, remains a mystery. Perhaps our ancestors survived the onslaught of these infections by developing resistance while those who did not simply died out.
Human HIVs include not only HIV-1 group M, the pandemic strain of HIV, but also HIV-1 strains N, O, and P, and HIV-2. We now know that all these viruses recently jumped from primates to humans in Africa, and it is probable that such transfers have occurred from time to time throughout our history, but remained unnoticed because they did not spread beyond the immediate area. It was the unique occurrence of HIV-1 group M spread from Africa to Haiti and on to the USA in the 1960s that prompted the first description of AIDS in 1980 and the isolation of the virus in 1983.
HIV-hospital-acquired virus2, discovered in 1986, is only 40% identical to HIV-1 and has a quite distinct origin, having been acquired from the sooty mangabey monkey in West Africa. Although this virus spreads in the same way, infects the same cell types as HIV-1, and also causes AIDS, it is less infectious than HIV-1 and has remained local to West Africa.
HIV-1 and AIDS Since humans have acquired HIV-1 only recently, we lack genetic resistance to the virus, and thus virtually every untreated infection eventually ends in death from AIDS. Just a few fortunate individuals are resistant to infection. Although AIDS was first described in gay men, and shortly afterwards injecting drug users and haemophiliacs were found to be at risk, worldwide the virus is mainly transmitted by heterosexual intercourse.
There are now 33 million people living with HIV, with around 2.7 million new infections, and 2 million deaths, per year. The virus has invaded virtually every country in the world, with the overwhelming impact in developing countries; 22 million people are living with HIV in sub-Saharan Africa. But even these startling figures belie the tragedy of the worst-hit African countries where life expectancy has tumbled to below 40 years by the wholesale death of previously healthy and productive adults, creating an economic downturn, severe poverty, and around 15 million AIDS orphans.
HIV infects cells bearing the CD4 marker, mainly helper T cells and tissue macrophages. Virus infection occurs through contact with the blood or genital secretions of a carrier, usually via a tear or abrasion in the epithelium lining the genital tract, or, commonly, an open sore caused by another sexually transmitted infection such as HSV, gonococcus, or syphilis. On entry, the virus initially targets Langerhans cells, the subset of macrophages that patrol the skin and epithelial surfaces, including the lining of the genital tract.
These cells then carry the virus to the local lymph glands, where literally millions of CD4 T cells congregate while taking a rest from circulating in the blood. Infection of these long-lived cells not only disseminates the virus throughout the body but also provides a site of persistence as the proviral genome integrates into their DNA.
The clinical course of HIV infection naturally divides into three stages: the acute, the asymptomatic, and the symptomatic phases, the last being manifest as AIDS. People infected with HIV often experience a primary illness known as the acute retroviral syndrome between one and six weeks after infection.
This is a fairly non-specific illness with fever, sore throat, swollen glands, a rash, and general aches and pains, and usually lasts up to 14 days followed by complete recovery. Initially, the virus multiplies freely in CD4 T cells, destroying over 30 million of them every day.
Levels of virus in the blood (called the viral load) rise to a peak in the first few weeks, after which the immune response kicks in, controlling but not completely clearing the virus. The viral load then falls, and by six months it has generally stabilized to a ‘set hospital-acquired virus’ level, the height of which depends on the strength of the immune response and is all important in predicting the further course of the disease; the higher the set point, the quicker the progression to AIDS.
In an untreated person, the asymptomatic phase of HIV infection lasts between 6 and 15 years depending on the viral set point, and although carriers in this phase are generally well, HIV continues its battle with their immune system, causing cumulative damage. Early on, the HIV genome in infected cells is fairly uniform, but the more it replicates, the more it throws up mutants, some of which can evade the immune response.
As these mutants prosper, an arms race develops between immune T cells and antibodies, on the one hand, and a series of immunity-evading virus mutants, on the other. CD4 T cells are pivotal to the continually evolving immune response, but HIV replicates in these cells and destroys them at such a rate that the body cannot keep pace. Eventually, the CD4 cell production line runs dry and numbers decline. Without antiviral drugs to control virus replication, the body’s capacity to replenish CD4 cells is eventually exhausted, such that when the level drops below the critical threshold of 200 CD4 cells per milliliter of blood, immunity to other pathogens fails and they take the opportunity to invade.
Evidence of declining immunity and the imminent onset of the symptomatic phase of the HIV infection, AIDS, often include weight loss, night sweats, recurrent chest infections, skin lesions such as warts, and oral ulcers and infections like thrush and cold sores. These are then followed by the relentless onslaught of a plethora of opportunistic infections, including reactivation of persistent microbes like CMV, HSV, VZV, and TB, as well as tumours caused by HPV, KSHV, and EBV. One of the hallmarks of AIDS is infection with microbes that are no problem to people with healthy immune systems, for example pneumonia caused by avian TB or the fungus Pneumocystic jirovecii, (previously P. carinii) – the latter provided the clue to the recognition of AIDS as a new disease in 1980.
Central nervous system manifestations are also common in AIDS, as HIV invades the brain at an early stage of the disease, infecting and killing cells, causing progressive degenerative changes leading to AIDS-associated encephalopathy and dementia. In addition, CMV and another very common, persistent, and generally asymptomatic virus called JC (from the initials of the patient from whom it was first isolated) may cause progressive degenerative brain disease in AIDS sufferers.
Death from one of these infections inevitably follows, often within months. Fortunately, today antiretroviral therapy has transformed this grim picture of HIV infection into a treatable chronic disease, but this treatment is not without its problems, and there are still millions of HIV sufferers in the developing world who have no access to these life-saving drugs.
Hepatitis viruses
Hepatitis, meaning inflammation of the liver, can be caused by a variety of viruses as well as toxic chemicals such as alcohol and the drug paracetamol. The liver is a huge organ with plenty of spare capacity, so mild inflammation often passes unnoticed. The main indication of more severe damage is the yellow discoloration of the skin known as jaundice, often most noticeable in the whites of the eyes.
Several viruses, including Epstein–Barr and herpes simplex viruses, can cause hepatitis as part of a generalized infection, but for others the liver is their main site of replication, causing them to be lumped together as ‘the hepatitis viruses’.
To date, five human hepatitis viruses have been discovered and named A, B, C, D, and E. With the exception of HDV, all these viruses either infect silently or produce clinical hepatitis varying in severity from mild and self-limiting to fulminate – that is, acute liver failure which is generally fatal unless a liver transplant can be performed as an emergency procedure.
Hepatitis A and E viruses spread by the faecal–oral route causing epidemics of ‘infectious jaundice’, and where standards of hygiene are low most children are infected at an early age. Although the illness may be prolonged, recovery is the rule, and the viruses do not persist thereafter. In contrast, hepatitis B and C viruses may persist after primary infection, and this can lead to chronic hepatitis, cirrhosis, and liver cancer. Hepatitis D virus (HDV), also known as delta virus, is unique among human viruses in being defective and requiring the assistance of HBV for its transmission.
Specifically, HDV particles consist of an RNA genome surrounded by their own protein but enveloped in HBV surface antigen that acts as its receptor for getting in and out of liver cells. So this virus can only replicate in cells already infected with HBV and manufacturing HBV surface antigen. HDV may be transmitted along with HBV or may infect an HBV carrier, and in both cases it tends to worsen the infection by increasing the liver damage and accelerating the onset of chronic liver disease.
Hepatitis C virus is mainly spread by blood contamination. Once routine testing of donor blood excluded most HBV-infected units in the 1970s, HCV became the commonest cause of viral hepatitis following blood transfusion. But after its discovery in 1989, when blood and blood products were screened for HCV, the commonest route of transmission became needle sharing by intravenous drug users. Around 10% of carrier mothers pass the virus to their newborn offspring, but household and sexual contacts are not thought to be at increased risk.
HCV presently infects around 170 million people. Infection occurs worldwide but shows marked geographical variation, with 1-2% of the population infected in the USA, northern Europe, and Australia, and rates of up to 5% in southern and central Europe, Japan, and parts of the Middle East. The highest levels of around 20% are recorded in Egypt, where a treatment programme for the parasitic disease bilharzia in the 1960s unwittingly spread the virus by using non-sterile needles.
Only about one-quarter of those with primary HCV infection develop hepatitis with symptoms, but whether symptomatic or not, around 80% of acute HCV cases progress to a chronic phase. HCV has many ways of dodging the body’s immunity. As an RNA virus, HCV, like HIV, mutates rapidly and this, combined with its extremely high replication rate, generates a whole array of minor genetic variants, called quasispecies, in a single individual. Some of these variants manage to evade immune T cells and antibodies generated specifically to combat the virus, and these mutants then flourish until the immune response catches up with them. Then another viral variant will come to
prominence, and this immune driven evolution will continue to foil host immunity ad infinitum. HCV also evades host immunity by blocking antiviral mechanisms inside infected cells, preventing the production of cytokines like interferon.
The virus also induces regulatory T cells that paradoxically damp down anti-HCV immunity. The importance of this is demonstrated by the finding that during primary HCV, the height of this response reflects the outcome: those with a high level of regulatory T cells have a higher viral load and are more likely to develop a persistent infection than those with a lower level of the same cells.
It is not clear whether the liver damage caused by HCV infection is directly due to virus replication in liver cells or to immunopathology, but whatever the mechanism, there are signs of ongoing liver damage in all chronic HCV carriers, many of whom are unaware of the infection, and this progresses to chronic active hepatitis and/or cirrhosis in up to 70% of cases. Intensive antiviral treatment can clear the virus in some cases, but this is expensive and only affordable by health services in the developed world.
No vaccine is available to prevent HCV infection, and with 3% of the world’s population currently infected, this is now the commonest cause of liver failure and indication for liver transplantation in the Western world. Chronic HCV infection is also associated with the development of liver cancer, and in countries where HBV prevalence has decreased due to the screening of donor blood and more recent vaccination programmes, HCV is now the major risk factor for this tumour. HBV was discovered by chance in 1964 in the blood of an Australian Aborigine and shown to be a major cause of transfusion-associated hepatitis. The virus is extremely infectious and carriers have high viral loads in blood and body fluids. It spreads by close contact, particularly sexual intercourse, and mother to child, as well as by blood contamination of medical instruments, dental drills, and needles used for injection, and household utensils such as razors, toothbrushes, and by tattooing, body piercing, and acupuncture. Intravenous drug users and gay men are at particular risk of infection. Around 350 million people worldwide carry HBV.


{kind=link}