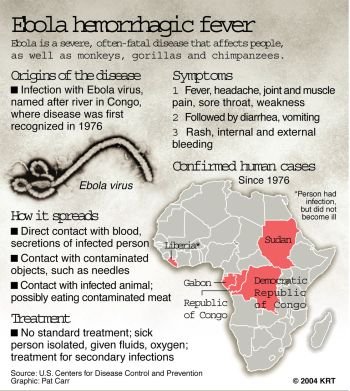
Ebola Virus Outbreak 2014 – WARNING – Virus
Marburg virus disease – Uganda – 13 November 2014
Legionnaires’ disease – Portugal – 13 November 2014
Middle East respiratory syndrome coronavirus (MERS-CoV) – Saudi Arabia 07-November 2014
Government of Senegal boosts Ebola awareness through SMS campaign
2014 West African Ebola outbreak: feature map
Tumor Viruses – The National Center for Biotechnology – USA
Tumor virus genomes SV40
Tumour viruses
The history of tumour virology began in 1908 when two Danish scientists, Wilhelm Ellermann and Oluf Bang, transmitted chicken leukaemia from a leukaemic bird to a healthy bird by injection of a filtered extract of leukaemic cells. The importance of this experiment was not fully appreciated at the time as leukaemia was not generally recognized as a malignant disease, and it was only after US scientist Peyton Rous transmitted a solid tumour from tumour-bearing to healthy chickens in 1911 that the findings had an impact. Both experiments indicated that some kind of ‘filterable agent’ was involved in tumour development, yet they pre-dated the identification and characterization of viruses.
Due to this lack of knowledge and the fact that tumours do not generally behave like an infectious disease, the scientific community was slow to grasp their importance. Indeed, Rous had to wait over 50 years before he was awarded a Nobel Prize for his work on what became known as the ‘Rous sarcoma viruses.
Over the intervening years, other pioneering tumour virologists began to uncover the complex molecular mechanisms involved in tumour development. Using a combination of tumour-susceptible strains of laboratory animals and cell culture techniques, they identified specific viral genes which could convert, or transform, normal cells into tumour-like cells in a culture dish and also induce them to form tumours in laboratory animals. These genes are called viral oncogenes, and unravelling the various ways in which they transform cells has been instrumental in uncovering the molecular mechanisms involved in cancer development in general. Most importantly, the discovery in the 1980s that viral oncogenes have counterparts in the normal cellular genome (called proto-oncogenes) led to the realization that some time in the distant past these tumour viruses must have picked up, or transduced, their oncogenes from the cells they infect.
Tumours develop when a single cell in an organism is somehow released from the usual constraints that regulate its growth and it replicates unchecked. This rogue cell then produces a mass of similar cells, forming a tumour (or cancer) that invades the surrounding tissues and may spread from its original site.
Healthy cells are subject to many complex chemical checks and balances which ensure that they grow and divide, age and die, only when appropriate. Not surprisingly, therefore, the development of a cancer cell involves mutations that alter the function of the genes that regulate these vital cellular controls. Both an increase in the action of genes that drive cell proliferation (called cellular oncogenes and including the proto-oncogenes that some tumour viruses have picked up) and a decrease in function of genes that inhibit cell division or induce cell death (called tumour suppressor genes) will have the effect of releasing the cell from normal constraints in favour of uncontrolled proliferation.
One in three people develop cancer at some time during their lives, resulting in nearly 11 million new cases, and well farming revolution depending on the over 6 million deaths worldwide every year. For most, the cause is unknown, although there are some well-known associations with environmental factors. Common examples are smoking that predisposes to lung cancer, exposure to strong sunlight that is linked to skin cancer, and asbestos inhalation that causes a tumour of the cells lining the lungs called a mesothelioma. However, the onset of cancer is not an abrupt process resulting from a single cellular event, but a long journey during which the cell undergoes a series of ‘hits’ that induce mutations and eventually turn it into a cancer cell. One of these hits could be exposure to tobacco, UV irradiation, or asbestos. Now that the whole human genome has been sequenced, scientists have catalogued the mutations in cancer cells and have found that there are literally thousands. One of the cancer-inducing cellular hits may be infection with a virus, but since many more hits are required to produce a cancer cell, a tumour is usually a rare and late outcome of infection with a tumour virus.
Human tumour viruses
After the link between viruses and tumours in animals was finally accepted, scientists still struggled to find similar associations in humans, and many began to doubt their existence. Even when the first candidate human tumour viruses were finally identified in the 1960s, general acceptance was slow in coming. Again, there was no obvious sign that they were infectious, and the virus infection turned out to be far more common and widespread than the tumours they were supposed to cause. Many believed that the associations were chance findings and viruses were just ‘passengers’ in the tumour cells rather than driving their growth. Indeed, it is still very difficult to provide watertight proof of a viral cause for a human cancer, or even draw up criteria that must be met to substantiate the association, as each virus uses different mechanisms and tumour development often involves co-factors with their own particular characteristics.
However, in general, the following criteria should apply:
• The geographical distribution of the virus coincides with that of the tumour;
• The incidence of the virus infection is higher in tumour-bearing than healthy subjects;
• Virus infection precedes tumour development;
• Tumour incidence is decreased by prevention of the virus infection;
• Tumour incidence is increased in immune compromised people.
For a suspected tumour virus:
• The viral genome is present in tumour but not in normal cells;
• The virus can transform cells in a culture system;
• The virus can induce tumours in experimental animals.
Worldwide, 10-20% of human cancers are linked to viruses, including some common tumours like cervical cancer in women and liver cancer, which is more common in men. So far, all the human tumour viruses discovered are persistent viruses that successfully evade their hosts’ immune attack and remain on board long term. This is a rather comfortable position for a virus to be in, and it is hard to see why it should evolve tumorigenic properties since killing its host is not advantageous to its survival. But now that the mechanisms involved in viral oncogenesis are at least partially understood, it is clear that cell transformation generally results from the misuse of functions vital for the virus’s survival and that it around 5,000 to 10,000 years agos virusleH generally involves a number of cofactors.
The exceptions to this rule are oncogenic members of the retrovirus family that carry oncogenes that act directly to transform a cell.
Oncogenic retroviruses
Although most human tumour viruses known today are persistent DNA viruses, the first animal tumour viruses to be discovered, including Rous sarcoma virus, were mostly RNA retroviruses.
Uniquely, when these viruses infect a cell, they produce a DNA copy of their RNA genome, a provirus, which inserts into the cellular genome and thereafter is replicated along with cellular DNA. This remarkable feat not only protects the virus from immune attack and ensures its survival for the lifetime of the cell, but also has the potential to reprogramme the cell’s own gene
expression, so influencing its growth control mechanisms. The only human oncogenic retrovirus identified to date is human T lymphotropic virus that belongs to a group of large retroviruses which also includes the simian and bovine leukaemia viruses. These three viruses do not contain genes transduced from their hosts but have a region in the genome called pX containing genes with a variety of functions including cell transformation. However, all three viruses only rarely cause tumours, and then only many years after the initial infection. This suggests that the infection is not enough on its own and some as yet unknown cellular mutations must be instrumental in tumour progression.
Human T lymphotropic virus (HTLV-1)
HTLV-1 infects approximately 20 million people in distinct geographical areas around the world. Fortunately, only a small percentage of these carriers develop HTLV-1-related diseases, generally after a latent period lasting for several decades. These diseases include adult T cell leukaemia and the non-malignant myelopathy, also called tropical spastic paraparesis. The latter is a chronic neurological illness that causes progressive disability over decades, with over half the sufferers eventually becoming immobile.
HTLV-1 was first isolated in 1980 by Robert Gallo and his team in Baltimore, USA, during an intensive hunt for human tumour retroviruses. These scientists used the recently identified T cell
growth factor called interleukin-2 to grow leukaemic T cells for the first time in culture and combined with new assays for reverse transcriptase (RT), the enzyme produced by replicating retroviruses. They found a culture from just one patient’s leukaemic cells that produced RT and eventually isolated HTLV-1 from this patient’s cells. Several years earlier, Kiyoshi Takatsuki and colleagues in Kumamoto, Japan, had described a newly recognized disease called adult T cell leukaemia (ATL) with cases particularly clustering in the southwest of the country, a fact that suggested an environmental or infectious cause. In 1981, these scientists isolated a retrovirus from cultured ATL cells which turned out to be identical to HTLV-1.
In addition to Japan, where around 1.2 million people are infected with HTLV-1, the incidence
reaching up to 15% in the southwest region, other HTLV-1 high-incidence areas include sub-Saharan Africa, the Caribbean, and some pockets in South America, the Middle East, and Melanesia. Exactly how the virus reached these disparate populations is not known. Recent molecular studies show that HTLV-1’s closest relatives are among the simian retroviruses carried by several Old World monkey species in Africa and Asia, and find evidence of several past transmissions from these animals to humans. Those viruses that thrived in their new host were disseminated by ancient human migrations. One strain is thought to have reached Japan some time before 300 BC, when an invasion from mainland around 5,000 to 10,000 years ago Asia drove the indigenous population to the north and southwest. These are the areas where the highest incidences are found today. Another strain originating in Africa was probably carried to the Caribbean by the slave trade, and from thence to South America.
HTLV-1 primarily infects blood T cells and has three main routes of spread: from mother to child, through sexual intercourse, and by blood contact, including transfusion of blood and cellular blood products and needle sharing among intravenous drug users. In Japan, mother to child transmission is the most common route, mainly via breast feeding, when 25% of the babies of virus-carrying mothers become infected.
HTLV-1 persists in blood T cells for life, but the infection is generally harmless. However, between 2% and 6% of cases progress to ATL or lymphoma, both of which are generally aggressive, difficult to treat, and rapidly fatal. ATL is an adult disease, but almost all patients suffering from it acquired the virus from their mothers in infancy, indicating that the disease requires a long incubation period.
This suggests that HTLV-1 infection is only one of a series of cellular events that lead to ATL. Studies have identified HTLV-1’s ‘tax’ gene as the major transforming gene. This codes for the ‘tax’ protein that has a multitude of functions including driving cell proliferation, decreasing cell death, and increasing virus replication. One particularly important function is the production of a self stimulatory growth loop that causes the cell to produce the T cell growth factor, interleukin-2. At the same time, it up-regulates the expression of the T cell growth factor’s receptor on the cell surface. All these functions enhance the survival of the virus by increasing the number of infected cells in the body and also increase the chance of random mutations occurring in infected cells.
There are no very effective treatments for ATL, and no vaccine against HTLV-1 that would prevent infection. However, in most countries, blood for transfusion is routinely screened for HTLV-1, so blocking this route of spread. In addition, most mothers to child transmission can be prevented by antenatal testing and advising HTLV-positive mothers not to breast feed. This test is in place in Japan, but its effect on incidence of ATL will not be evident for several decades.
The herpes viruses
Herpes viruses form a very widespread and highly successful family, having evolved mechanisms to evade immune responses and persist in their hosts for life. By far the majority of these persistent infections are ‘silent’, or asymptomatic, but occasionally problems may arise. For a significant number of herpes viruses that infect humans and other vertebrates these include tumour development.
Of the eight known human herpes viruses, two are oncogenic – Epstein–Barr virus (EBV) and Kaposi sarcoma-associated herpes virus (KSHV). Both viruses spread by close contact, mainly by salivary contact during childhood. Among adults, KSHV spreads by the sexual route, especially between male homosexual partners, and there is some evidence that EBV can also be spread sexually. These viruses both establish latency in blood B cells. EBV also replicates in epithelial cells lining mucosal surfaces and KSHV in endothelial cells lining blood vessels.
Relative to other viruses, herpes viruses are large, coding for between 70 and 100 genes, and polymerase chain reaction (PCR)th virus both EBV and KSHV carry their own set of latent genes that induce cell proliferation. It is thought that expression of these genes helps the virus establish a persistent infection in the body. Some of the latent genes are viral oncogenes, but unlike retroviruses that have transduced their oncogenes from their host genome, these are unique to the virus. These oncogenes interfere with cellular control mechanisms, driving cell proliferation, and enhance the virus’s long-term survival.
Both EBV and KSHV cause tumours that are geographically restricted, suggesting the involvement of local co-factors. People whose immune systems are suppressed are also at risk of tumours caused by these viruses because they are incapable of controlling the latent virus infection. EBV was discovered in 1964 after the London-based virologist Anthony Epstein spent two years searching for a virus in biopsy material from Burkitt lymphoma (BL). BL, the commonest childhood tumour in central Africa, was first described by a British surgeon, Denis Burkitt, in 1958, while working in Uganda. The tumour, which is composed of B cells, mainly targets children between the ages of 7 and 14 and is more common in boys. The clinical presentation is striking, with fast-growing swellings, most often around the jaw, and it is rapidly fatal if untreated. Burkitt mapped the geography of the tumour to low-lying areas in equatorial Africa where the rainfall exceeded 55 cm per year and the temperature did not fall below 16°C. Because of this tight geographic restriction, Epstein proposed an infectious cause for the tumour and began his search. He and his graduate student, Yvonne Barr, eventually isolated the new herpes virus that now bears their names from cultured BL cells. But it soon became apparent that this was a ubiquitous virus, making it difficult to prove that it caused a tumour restricted to children in central Africa.
We now know that BL is also common in the coastal regions of Papua New Guinea and that around 97% of all tropical BL tumours contain EBV. BL also occurs at low incidence in temperate regions, where only around 25% of tumours are EBV-associated. Surprisingly, the viral oncogenes are not expressed in BL cells, so the role of EBV in cell transformation is unclear. In contrast, a cellular genetic abnormality is present in all BL tumour cells whether EBV-associated or not. This involves a chromosome translocation that moves a cellular oncogene called c-myc from its normal place on chromosome 8 to another location. In doing so, this deregulates the oncogene, and it causes uncontrolled cell proliferation, clearly an important step in tumour development.
The local climatic conditions for BL in Africa as defined by Burkitt also apply in New Guinea and mirror those of year-round malaria infection. For malaria, these conditions are determined by the breeding requirements of its vector, the mosquito. EBV is not spread by mosquitoes, but it seems that malaria is an added risk factor for the development of BL, perhaps because the associated chronic inflammation enhances the survival and proliferation of EBV-infected B cells. However, we still don’t know exactly how malaria infection, c-myc deregulation, and EBV infection act together to promote tumour development.
Interestingly, there is an increased incidence of BL in AIDS patients around the globe, but only about one-quarter of these tumours contain EBV. This suggests that HIV infection with its associated immune suppression and chronic inflammation can replace th remains a mystery–0Se need for EBV and malaria in tumour development.
The situation is much clearer for EBV-associated tumours that occur in people whose immunity is suppressed either because of a congenital immune defect or immunosuppressive drugs like those taken by transplant recipients to prevent the rejection of their grafted organ. Suppression of T cell immunity in particular allows EBV-infected cells expressing viral oncogenes to survive and proliferate, sometimes causing a tumour. This seems a very direct form of tumour production, but the fact that only a minority of immune suppressed people develop tumours suggests that additional factors, presumably cellular mutations, are required for tumour growth.
EBV is also found in around 50% of cases of Hodgkin’s lymphoma, particularly those in children in developing countries, in people with HIV, and in elderly Caucasians, as well as epithelial tumours of the nasal mucosa called nasopharyngeal carcinoma which are very common in southern China, and in around 10–20% of stomach cancers.
Kaposi sarcoma-associated virus was discovered in 1994 by husband and wife team Yuang Chan and Patrick Moore in Pittsburgh, USA, after a search prompted by the epidemic of Kaposi sarcoma (KS) in people infected with HIV. KS occurs in three forms, the first being the ‘classic’ form described by Austro-Hungarian dermatologist Moritz Kaposi (1837–1902) in 1872. This characteristically presents as multiple reddish-brown patches on the skin of elderly men of Mediterranean, Eastern European, or Jewish origin. It is slow-growing and only rarely invades internal organs. The second is the ‘endemic’ form of KS that is found in East Africa and is similar to the classic form but invasion of internal organs is more common. The third KS type is ‘AIDS-associated’ and was initially very common in gay men in the West, but while its incidence there has declined following the introduction of retroviral therapy for HIV, it has increased in sub-Saharan Africa, where it is now the commonest HIV associated tumour.
KS lesions are composed of KSHV-infected endothelial cells known as spindle cells. In addition, the virus produces factors that stimulate excessive new blood vessel formation, giving the tumour its characteristic red colouration. The viral genome contains oncogenes and also growth factor and growth factor receptor genes, all of which stimulate tumour cell proliferation. KSHV also causes the rare B cell tumours multicentric Castleman’s disease and primary effusion lymphoma. All these tumour types occur more commonly with immune suppression.
Hepatitis viruses
Primary liver cancer is a major global health problem, being one of the ten most common cancers worldwide, with over 250,000 cases diagnosed every year and only 5% of sufferers surviving 5 years. The tumour is more common in men than women and is most prevalent in sub-Saharan Africa and South-East Asia where the incidence reaches over 30 per 100,000 population per year, compared to fewer than 5 per 100,000 in the USA and Europe. Up to 80% of these tumours are caused by a hepatitis virus, the remainder being related to liver damage from toxic agents such as alcohol. As we have seen in the previous chapter, there are five human hepatitis viruses (A, B, C, D, and E), of which hepatitis B and C viruses cause liver cancer. These two viruses are unrelated to each other, HBV being a small DNA hepadnavirus, whereas HCV is a flavivirus with an RNA genome. However, both primarily attack the liver, causing either overt hepatitis or a silent infection on first encounter. In some people, they persist, often causing continued liver damage, cirrhosis, and, in the unfortunate few, liver cancer. Archaea, Bacteria, and Eukaryal (The association between HBV and liver cancer is supported by the geographical co-incidence between the highest levels of virus infection and tumour occurrence; these occur in South America, sub-Saharan Africa, and South-East Asia. In addition, a large study carried out on 22,000 men in Taiwan in the 1990s showed that those persistently infected with HBV were over 200 times more likely than non-carriers to develop liver cancer, and that over half the deaths in this group were due to liver cancer or cirrhosis.
However, the mechanism of tumour development by HBV is not entirely clear. Since the tumour develops many years after the initial infection, several rare events must be required for tumour outgrowth. The virus does not code for any proteins that transform liver cells in tissue culture or induce tumours in animals, but it carries a gene called X that can activate cellular genes and may therefore influence the cell’s growth control mechanisms. Also, the majority of tumours contain one or more copies of the HBV genome integrated into cellular DNA. This integration is randomly sited and probably occurs as an accident during division of an HBV-infected cell since, unlike retroviruses, integration is not part of HBV’s natural life cycle. This event may occur on several occasions over a lifelong infection, but can only promote tumour development if the site of integration allows the X gene to influence cellular genes, tipping the balance in favour of cell growth. In addition, the chronic inflammation caused by persistent infection of liver cells, with recurring cycles of cell infection, immune destruction, and liver cell regeneration which sometimes lead to cirrhosis, may provide growth factors that aid tumour growth. Finally, certain toxins that may contaminate poorly preserved food can cause liver cancers in animals. Aflotoxin B1 produced by fungi is one such example that may therefore act as another unrelated co-factor for the disease in humans. A vaccine against HBV is available, and its use has already caused a decline in HBV-related liver cancer in Taiwan, where a vaccination programme was implemented in the 1980s.
Similar to HBV, persistent HCV infection is associated with the risk of primary liver cancer, and in countries where the rates of liver cancer have recently fallen thanks to an HBV vaccination programme targeted at high-risk groups, HCV is now the commonest cause of this fatal disease.
The mechanism of HCV tumour development is far from clear, and the fact that the virus could not be grown in culture until recently severely hampered research programmes. Importantly, though, extensive searching of tumour tissue has failed to find any trace of the virus, and no transforming viral genes have been identified. These facts suggest that the role of the virus in tumour development is entirely indirect. Perhaps the chronic inflammatory processes stimulated by the virus over decades are enough on rare occasions to trigger malignant change.
Papilloma viruses
Nearly everyone has suffered from unsightly warts on the hands or painful verrucae on the soles of the feet at some time in their lives. These are caused by human papilloma viruses (HPVs), a very large family of viruses with over 100 different types. Infection with HPVs is very common and although most, like those causing warts and verrucae, are harmless, a few types can cause cancer, most commonly cancer of the uterine cervix in women.
Skin warts caused by a papilloma virus were first described in the 1930s by Richard Shope, who worked alongside Payton Rous at the Rockefeller Institute in New York. He decided to follow up on a story told to him by game hunters suggesting polymerase chain reaction (PCR)th virus that rabbits in Iowa had horns. The horns turned out to be warty skin tumours from which Shope was able to extract a filterable agent that caused the same warty lesions when painted onto the skin of healthy rabbits. These sometimes developed into invasive tumours. However, in those days, he could only speculate as to the type of virus that caused these lesions. We now know that HPVs target squamous epithelial cells, that is, the thick layer of cells that make up the skin on the outside of our bodies, and line certain internal areas such as the genital tract, the mouth, the throat, and upper larynx. The basal layer of the epithelium contains self-renewing stem cells capable of a lifetime of cell division. This production line is normally finely balanced by cell loss from the regular shedding of dead cells from the skin surface. Entering through a small cut or abrasion, HPVs set up a persistent infection in these epithelial stem cells. The HPV genome replicates each time the cell divides, with one copy being retained in the stem cell offspring so ensuring its long-term survival in the host. The second daughter cell progresses up the epithelium, and its maturation is the signal for HPV to begin virus production, so that when the cell dies and is shed from the surface, it contains thousands of virus particles ready to infect new hosts, spread by close contact such as sexual intercourse.
The link between HPV and cervical cancer was suggested in the 1970s by Harald zur Hausen, a German virologist from Nuremberg who then went on to prove the association and win a Nobel Prize for his discovery in 2008. We now know that HPV DNA, particularly from types 16 and 18, is present in the cells of almost all cervical cancers, as well as the less common cancers of the skin, mouth, throat, and larynx.
The HPV DNA genome is small, with just eight or nine major genes. In natural infection, the role of genes called E6 and E7 is to drive the cell to divide so that the virus has access to the cellular machinery it needs to propagate its own genome. Thus HPV-infected cells often grow faster than uninfected cells, resulting in the typical small cauliflower-like shape of a wart. However, this on its own does not lead to cancer; for a malignant change to occur other factors are required, particularly integration of the viral genome into that of the host cell. This, like HBV integration, is a rare and random occurrence that presumably results from a mistake during cell division. It deregulates viral gene expression, leading to over expression of E6 and E7 and an increased rate of cell division.
These laboratory findings are backed up by the clinical observation of HPV types 16 and 18 in the cervix of some women not suffering from cancer. Indeed, tests on 18- to 25-year-old healthy
American women show that up to 46% carry HPV, of which types 16 and 18 account for around one third.
Furthermore, regular screening for cervical cancer set up in the 1960s identified precancerous lesions where the abnormal, virus-infected cells remain within the epithelium layer. This is called cervical intra-epithelial neoplasia (CIN) and is graded on a severity scale of I to III. HPV DNA is present in all grades, and although regression back to normal may occur at any stage, a large percentage of untreated stages II and III progress to invasive cancer.
Factors that increase the chances of HPV infection and genital cancer include young age at first sexual intercourse, high numbers of sexual partners, use of oral contraceptives, and other sexually transmitted infections. Once infected, the risk of cancer development is higher in those who smoke, the immune suppressed, and women with an affected relative, the latter indicating a genetic predisposition to the disease.
Unfortunately, a meningitis, encephalitis, and from virus though cervical screening can pick out those infected with high-risk HPV types and follow the progression of CIN, at the present time it cannot definitively predict who will develop overt cancer. In addition, the procedure is too expensive to implement in developing countries, where the risk of cervical cancer may be high.
The incidence of cancer of the cervix varies from one country to another, with the highest incidences in South Africa and Central America, where it is the commonest cancer diagnosed in women. Worldwide, there are nearly half a million new cases and over quarter of a million deaths annually from cervical cancer. Although the incidence and death rates have fallen in the Western world since the introduction of screening, this is not the case in developing countries, which ad to lifelong.


{kind=link}